Publication
Article
Oncology Live®
TIGIT Emerges as New Target for Immune Checkpoint Blockade Strategies
Author(s):
As researchers continue to identify a growing number of immune checkpoints as targets for anticancer therapy, the recently discovered TIGIT pathway is emerging as a promising new avenue for exploration.


As researchers continue to identify a growing number of immune checkpoints as targets for anticancer therapy, the recently discovered TIGIT pathway is emerging as a promising new avenue for exploration.
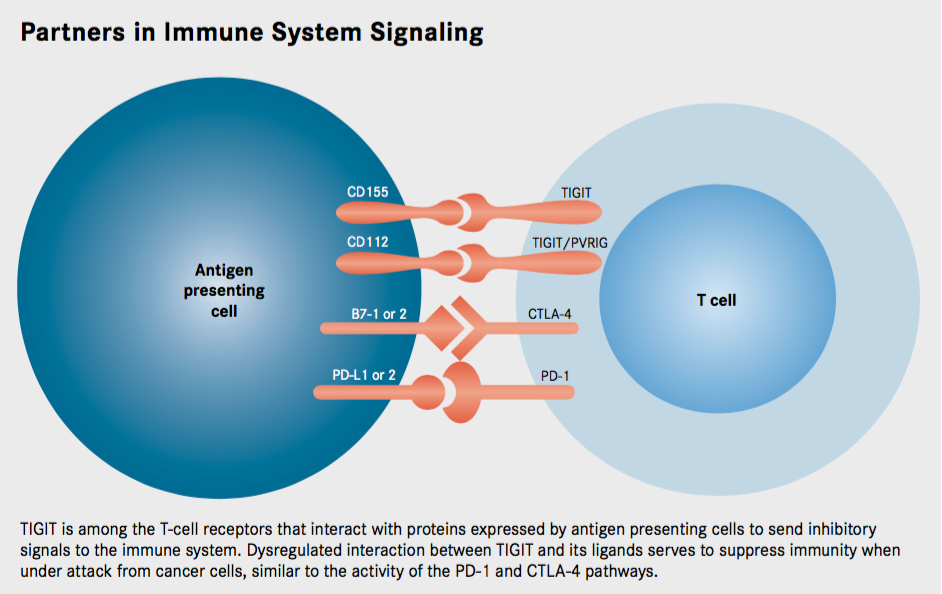
TIGIT is a poliovirus receptor (PVR)—like protein, an immunoreceptor expressed on T cells that contains immunoglobulin (Ig) and immunoreceptor tyrosine–based inhibitory motif (ITIM) domains. As such, TIGIT acts as an inhibitory immune checkpoint on both T cells and natural killer (NK) cells, providing an opportunity to target both the adaptive and innate arms of the immune system.
Although clinical development is in preliminary stages, TIGIT and related proteins show significant therapeutic promise, particularly in combination with other immune checkpoint inhibitors, with the potential to broaden the benefit of immunotherapy into previously unresponsive patient populations.
Genentech has launched a phase I trial evaluating MTIG7192A, a fully human monoclonal anti- body that binds to TIGIT and prevents its interaction with PVR. The trial, which opened in June, will evaluate the safety and efficacy of MTIG7192A as monotherapy and in combination with the PD-L1 inhibitor atezolizumab (Tecentriq) in a 2-step study that aims to enroll 300 patients with locally advanced or metastatic tumors (NCT02794571).
In September 2016, Bristol-Myers Squibb initiated a phase I/II study of an anti-TIGIT monoclonal antibody, BMS-986207, as monotherapy and in combination with nivolumab (Opdivo) in advanced solid tumors (NCT02913313). The estimated enrollment for the trial is 170 participants.
Where TIGIT Fits In
Compugen Ltd is developing COM701, an antibody that targets PVRIG, another recently elucidated member of the PVR family. The company is planning to submit an Investigational New Drug application to the FDA during the fourth quarter of 2017.The major effectors of the immune system are cytotoxic T cells, and these are activated in a 2-step process when they encounter antigen-presenting cells bearing foreign antigens. Antigens engage the T-cell receptor (TCR) on the surface of the T cell, but this signal is coupled with a second signal that determines whether the T cell is turned on or off. Together these costimulatory and coinhibitory signals are known as immune checkpoints.
CTLA-4 and PD-1, the most successful immune targets for anticancer therapies, generate coinhibitory signals. Since cancer cells and the cells of the surrounding microenvironment have been shown to upregulate the expression of components of their respective pathways as a means of suppressing the antitumor immune response, the development of antibodies that block their activity has been embraced by immuno-oncologists in the hopes of reversing these suppressive effects.
The anti—PD-1 antibodies nivolumab and pembrolizumab (Keytruda) and the PD-L1–targeting agent atezolizumab have been approved in multiple malignancies while ipilimumab (Yervoy) remains the only CTLA-4 inhibitor on the market.
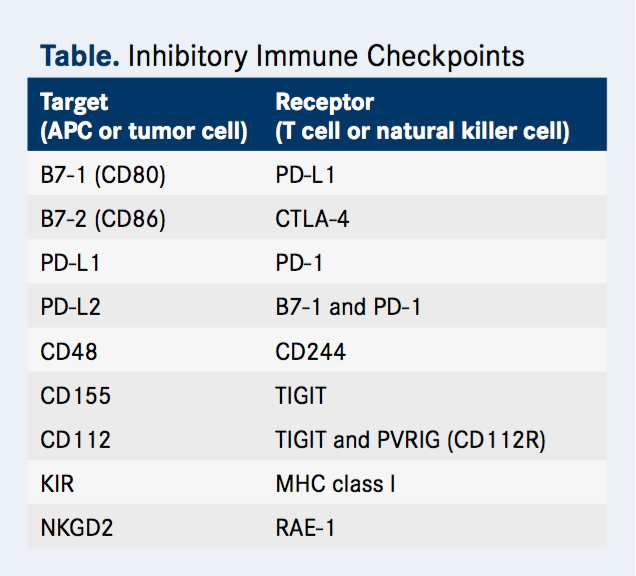
TIGIT is among the novel coinhibitory immune checkpoints under study (Table). It was first identified about a decade ago in a genome-wide screen- ing for potential immune inhibitory proteins. Researchers were searching for proteins that were expressed by immune cells and that contained an ITIM domain, which is known to mediate immune cell deactivating signals. TIGIT is a member of a recently discovered arm of the Ig superfamily, the PVR-like proteins, which contain PVR motifs in their Ig variable-like domain.
Further investigation revealed that TIGIT was reminiscent of the CTLA-4 protein, in that it shares ligands with an activating receptor. CTLA-4 is activated by binding to the B7-1 and B7-2 (also known as CD80 and CD86, respectively) proteins. These proteins also serve as ligands for the CD28 protein, a costimulatory molecule; thus the activating and deactivating receptors compete for the same ligand, with a delicate balance determining if the T cell is switched on or off.
The ligand for TIGIT is CD155 (alternatively known as PVR), but this protein also serves as a ligand for CD226, which, like CD28, is an activating receptor. When CD155 is bound to CD226, it conveys activating signals into the immune cell. Meanwhile, CD155 bound to TIGIT transmits an inhibitory signal by recruiting the SHP1 phosphatase to the membrane through its ITIM domain that subsequently deactivates numerous proteins involved in T-cell effector functions.
A third member of the PVR-like protein family, CD96 (sometimes referred to as T cell-activated increased late expression [TACTILE]), also binds to CD155. The function of this interaction is not yet understood, although mouse CD96 has been shown to be involved in inhibitory signaling like TIGIT.
Meanwhile, although TIGIT binds most strongly to the CD155 protein, it also partners with another ligand, CD112, with lower affinity, adding to the complexity of this pathway. Furthermore, PVRIG (known as CD112R), another newly discovered member of the PVR-like family, also binds to CD112.
TIGIT is expressed by activated cytotoxic T cells and regulatory T cells and has also been shown to be upregulated on T cells in multiple cancer models. The ligands CD155 and CD112 are found on dendritic cells and macrophages and are also highly expressed in several types of cancer. Additionally, TIGIT expression is highly correlated with the expression of other coinhibitory molecules, including PD-1. Overall, this suggests that tumors upregulate the TIGIT pathway along with other inhibitory checkpoint networks to promote immunosuppressive mechanisms.


Case 1 - BRAF Wild-type Stage IV Melanoma
Ways to Attack TIGIT
Thus far, the development of drugs targeting TIGIT and other PVR-like proteins remains in the preliminary stages, but if they live up to the promise of preclinical studies they could prove to be an important addition to the immunotherapy arsenal.
TIGIT-targeting antibodies are the furthest along in development, although only a few agents have progressed to clinical trials thus far. Such antibodies disrupt binding of TIGIT to its ligands and block its inhibitory signals, shifting the balance in favor of CD226-mediated activating signals. Preclinical studies demonstrated that these drugs enhance the function of T cells and inhibit the growth of tumors, particularly when combined with blockade of other inhibitory checkpoints, such as PD-1.
In a preclinical study presented at the 2016 Annual Meeting of the Society for Immunotherapy of Cancer in November, the combination of TIGIT and PD-L1 blockade in humanized mouse models led to enhanced CD8-positive T-cell function, which subsequently significantly improved tumor clearance. The activity of TIGIT was dependent on the expression of its ligand, CD155, in the host tissue.
A Unique Checkpoint
In data presented about the molecule COM701, PVRIG blockade resulted in enhanced activation of tumor-infiltrating lymphocytes, CD4-positive T cells, and CD8-positive T cells derived from the tumor. The use of a surrogate antibody, with similar characteristics to COM-701, showed synergy with anti-PD-L1 blockade in vivo in multiple cancer models. Compugen also reports on its website that in vitro studies have shown that dual blockade of PVRIG and TIGIT increases the activity of tumor-infiltrating T cells above the levels achieved by monotherapy against either target.Several studies have revealed TIGIT as a unique and potentially complementary target to other inhibitory immune checkpoints. First, in addition to directly inhibiting cytotoxic T-cell activity, TIGIT can foster an immunosuppressive microenvironment through its impact on other immune cells. For example, by binding to CD155 on the surface of dendritic cells, TIGIT increases the secretion of the immunosuppressive cytokine interleukin-10, and engagement of TIGIT on regulatory T cells enhances their immunosuppressive functions.
Second, TIGIT is also expressed on NK cells, the principal effector of the innate immune response that has been gaining traction as a target for anti-cancer therapy in recent years. Immune checkpoints have also been identified on NK cells, such as killer immunoglobulin-like receptor (KIR) that can be targeted to manipulate NK cell activity. In preclinical models, TIGIT blockade also boosts NK cell activation, suggesting that targeting TIGIT could offer a way of simultaneously boosting both arms of the immune response.
Most intriguingly, TIGIT has been shown to not simply outcompete CD226 for binding to CD155, but also to physically impede the dimerization of the activating receptor, blocking its costimulatory function. Thus, TIGIT inhibitors might not only release the brakes on the immune system, but at the same time may hit the gas by releasing TIGIT inhibition of CD226.
Certainly, the complexities of TIGIT and CD226 signaling are posing opportunities and challenges for clinical translation, and a fundamental understanding of these pathways will be essential going forward.
REFERENCES
Blake SJ, Dougall WC, Miles JJ, et al. Molecular pathways: targeting CD96 and TIGIT for cancer immunotherapy. Clin Cancer Res. 2016;22(21):5183-5188. doi:10.1158/1078-0432.CCR-16-0933.
Grogan J, Manieri N, Chiang E, et al. The immunoreceptor TIGIT regulates anti-tumor immunity. J Immunother Cancer. 2016:4(suppl 1):P209.
Kurtulus S, Sakuishi K, Ngiow SF, et al. TIGIT predominantly regulates the immune response via regulatory T cells. J Clin Invest. 2015;125(11):4053- 4062. doi:10.1172/JCI81187.
Levy O, Chan C, Cojocaru G, et al. Computational identification, functional characterization, and antibody blockade of a new immune checkpoint in the TIGIT family of interacting molecules. Presented at: 2016 SITC Annual Meeting; November 9-13, 2016; National Harbor, MD. Abstract O3.
Lozano E, Dominguez-Villar M, Kuchroo V, Hafler DA. The TIGIT/CD226 axis regulates human T cell function. J Immunol. 2012;188(8):3869-3875. doi:10.4049/jimmunol.1103627.
Mahnke K, Enk AH. TIGIT-CD155 Interactions in melanoma: a novel co-inhibitory pathway with potential for clinical intervention. J Invest Dermatol. 2016;136(1):9-11. doi:10.1016/j.jid.2015.10.048.
Pauken KE, Wherry EJ. TIGIT and CD226: tipping the balance between co-stimulatory and coinhibitory molecules to augment the cancer immunotherapy toolkit. Cancer Cell. 2014;26(6):785-787. doi:10.1016/j.ccell.2014.11.016.
Stanietsky N, Simic H, Arapovic J, et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc Natl Acad Sci U S A. 2009;106(42):17858-17863. doi:10.1073/pnas.0903474106.
Zhu X, Paniccia A, Schulick AC, et al. Identification of CD112R as a novel checkpoint for human T cells. J Exp Med. 2016;213(2):167-176. doi:10.1084/jem.20150785.
























